|
بوم شناسی بوم شناسی علم مطالعه روابط موجودات زنده با همدیگر و همچنین محیط آنهاست
| ||
|
لینک دوستان
|
مقدمه: مکانیسم جذب یون ها توسط سلول های گیاهی طی سال های اخیر مورد مطالعات دقیق قرار گرفته است. علی رغم آنکه ارتباط یونی بین سلول های گیاهی با محیط اطراف از چندین نظر با سلولهای جانوری اختلاف دارد، اما به طور کلی می توان از مطالب مربوط به الکتروفیزیولوژی سلول های جانوری و ویژگی های سلول های عصبی، برای سلول های گیاهی نیز استفاده نمود. از طرف دیگر کوچک بودن اندازه سلول های گیاهی، کار مطالعه آنها را مشکل می سازد. بنابراین دانشمندان اغلب در مطالعات خود از سلول های بزرگ جلبک های مختلف که دارای هستۀ مرکزی می باشند، استفاده می نمایند و شاید به همین دلیل اطلاعات در مورد جلبک ها از گیاهان بیشتر است. منبع پایگاه اینترنتی دکتر قلعه نویی موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ دو شنبه 20 شهريور 1391برچسب:فرآیند جذب در گیاهان, ] [ 9:54 ] [ حمیدرضا قاسمی ]
[ دو شنبه 9 مرداد 1391برچسب:فرم نحوه امتیاز دهی مرحله مصاحبه دکتری سراسری 91, ] [ 10:52 ] [ حمیدرضا قاسمی ]
گیاهان که منبع غذاها ، داروها و تعداد بیشماری از مواد آلی گوناگون هستند، در حقیقت گنجینهای عظیم از ثروت پنهانی بشمار میروند که پیوسته تجدید میشوند. گیاهان علاوه بر آنکه نقش تلمبه آب بیاندازه پرتوانی را میان خاک و جو ایفا میکنند. با بقایای فسیلی خود منشا منابع لازم برای تمدن کنونی هستند. سلول گیاهی آزمایشگاه بنیادی این کارخانه شگرف ترکیبات آلی است. مهم آن است که تعیین شود گیاه با چه فرآیندهایی (فتوسنتز، تعرق و (واکنشهای متابولیسمی|متابولیسم)) دگرگونیهای متعددی را باعث میشود که از چند ماده ساده آغاز میشوند و به تعداد بیشماری از پیچیدهترین مواد آلی حاصل از متابولیسم گیاهی میرسند.
منبع: سایت بی بوک موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ جمعه 6 مرداد 1391برچسب:بیوشیمی گیاهی, ] [ 12:41 ] [ حمیدرضا قاسمی ]
فیزیولوژی دانشی است که وظیفهاش بررسی عملکرد (Function) موجودات زنده است. ماهیت بررسی در این علم ، وظیفه و کارکرد اندامهاست. نام قدیمی فیزیولوژی وظایفالاعضا بوده است. فیزیولوژی گیاهی ، مطالعه اعمال حیاتی گیاه ، فرایندهای رشد و نمو ، متابولیزم و تولید مثل گیاهان است. منبع:
سایت بی بوک
موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ جمعه 6 مرداد 1391برچسب:فیزیولوژی گیاهی, ] [ 12:31 ] [ حمیدرضا قاسمی ]
برای دانلود سرفصل های دروس مقطع دکتری زیست شناسی گیاهی، روی نام آن ها کلیک کنید. منبع: http://www.sbu.ac.ir/Default.aspx?tabid=5150 موضوعات مرتبط: اکولوژی، تکوین گیاهی، فیزیولوژِی گیاهی، سیستماتیک گیاهی، ، برچسبها: [ پنج شنبه 5 مرداد 1391برچسب:سرفصل های دروس مقطع دکتری زیست شناسی - فیزیولوژی گیاهی, سلولی و تکوین گیاهی و سیستماتیک گیاهی, ] [ 23:46 ] [ حمیدرضا قاسمی ]
مکانیزمهای فعال و غیرفعال پیشنهادی جهت توجیه انتقال در آوند آبکش نظریات مرتبط با انتقال در آوند آبکش را می توان به دو دسته فعال و غیرفعال تقسیم بندی کرد. فرض هر دو نوع نظریه بر این است که بارگیری در منبع و جذب در مخزن مستلزم صرف انرژی است با این تفاوت که نظریات فعال معتقد به صرف انرژی برای فرایند انتقال هستند و حال آنکه نظریات غیرفعال بر این باورند که انرژی، تنها برای حفظ کارایی عناصر غربالی و نه برای تحریک فرایند انتقال لازم است. یکی از اصلی ترین نظریات فعال آن است که مواد محلول موجود در یک سوی عنصر غربالی، به کمک جریانهای سیتوپلاسمی و حرکتهای دورانی پروتوپلاسم، به سوی دیگر آن حرکت کرده و از طریق برخی مکانیزمهای ناشناخته از خلال صفحه غربالی عبور می کنند. از آنجا که هرگز جریانهای سیتوپلاسمی در عناصر غربالی بالغ و درحال کار رویت نشده است لذا باید این نظریه را مردود دانست. افزون بر این هیچ شاهدی مبنی بر حضور رشته های نازک اکتین، که در عمل جریانهای سیتوپلاسمی در سایر سلولهای گیاهی دخیلند وجود ندارد. مدل فعال اولیه دیگر برای انتقال در آوند آبکش بیانگر آن است که پروتئین P، همانند رشته های نازک اکتین در ماهیچه ها و میکروتوبولها در مژکداران و تاژکداران، قادر است از طریق برخی حرکات دودی و انقباضی، نیروی محرکه لازم برای حرکت مواد محلول را فراهم سازد، هر چند که شواهدی مبنی بر شباهت پروتئینP به اکتین یا توبولین وجود ندارد.
نوع دوم نظریه فعال بر این اعتقاد است که انرژی سبب می شود که مواد محلول از طریق صفحه غربالی، از یک عنصر غربالی به عنصر دیگر راه یابند. براساس تصاویر الکترونی بافت آبکش، این گونه به نظر می رسد که منافذ صفحه غربالی عموماً با کالوز و پروتئین P مسدود می شوند، لذا وجود مکانیزم انتقال فعال مواد محلول الزامی است. اگر به هنگام کار، منافذ صفحه غربالی واقع در بین دو عنصر غربالی بسته شوند، آن گاه مدلهای غیرفعال انتقال بر مبنای جریان توده ای مواد محلول، غیرممکن خواهد بود. اما انسداد ظاهری منافذ صفحه غربالی در این تصاویر، مصنوعی و ناشی از روش تثبیت نمونه می باشد به هنگام کار، منافذ صفحه غربالی واقعاً بازند و بنابراین وجود مکانیزم انتقال فعال مواد حل شونده از صفحات غربالی ضرورتی ندارد.
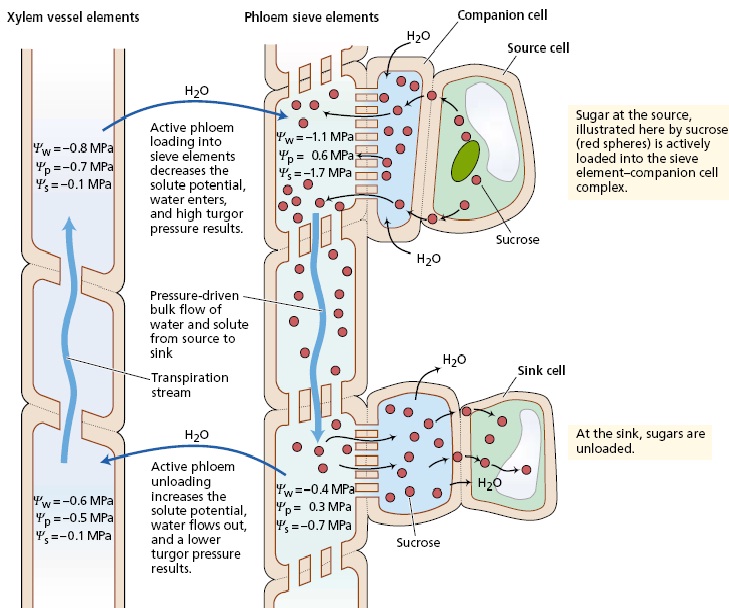
براساس فرضیه جریان فشار، انتقال در آوند آبکش به کمک شیب فشار از منبع به مخزن انجام می گیرد نظریات غیرفعال انتقال، فرضیات انتشار و جریان فشار را شامل می شوند. انتشار کندتر از آن است که بتوان روی سرعت حرکت مواد محلول مشاهده شده در آوند آبکش تاکید کرد سرعت انتقال در آوند آبکش 1 متر در ساعت است، حال آنکه سرعت انتشار 1 متر در 8 سال می باشد! از سوی دیگر، فرضیه جریان فشار به عنوان محتمل ترین مکانیزم انتقال در آوند آبکش پذیرفته شده است این فرضیه نخستین بار به وسیله مانچ در سال 1930 ارائه شده و بیانگر آن است که جریان محلول در عناصر غربالی، تحت اثر اختلاف شیب فشار بین مخزن و منبع صورت می گیرد. اختلاف فشار به دلیل انجام بارگیری در منبع و تخلیه در مخزن تداوم می یابد. بارگیری آوند آبکش سبب افزایش فشار اسمزی عناصر غربالی در بافت منبع شده و این امر افت شدید پتانسیل آب را در پی دارد. در پاسخ به این شیب پتانسیل آب آب وارد عناصر غربالی می شود و فشار تورژسانس آن را افزایش می دهد. در انتهای مسیر انتقال، تخلیه موجب کاهش فشار اسمزی عناصر غربالی در بافت مخزن می شود. همزمان با افزایش پتانسیل آب آوند آبکش و برتری آن بر آوند چوبی، آب در پاسخ به شیب پتانسیل آب، آوند آبکش را ترک می کند و این امر، کاهش فشار تورژسانس عناصر غربالی آبکش مخزن را در پی دارد. اگر در طول مسیر انتقال هیچ دیواره ای وجود نداشت (که وجود دارد) و اگر تمام مسیر انتقال یک جزء غشایی واحد بود آنگاه اختلاف فشار منبع و مخزن به سرعت به تعادل می رسید، اما وجود صفحات غربالی مقاومت مسیر را به شدت افزایش می دهد و منجر به تولید و حفظ اختلاف فشار بین عناصر غربالی منبع و مخزن می شود. از دیدگاه فیزیکی، جریان توده ای درست همانند حرکت آب از درون یک لوله آبرسانی، محتویات عناصر غربالی را در طول مسیر انتقال به جلو می راند. توجه کنید که براساس این مدل، در سرتاسر گیاه مقداری از آب بین مسیرهای تعرق (آوند چوبی) و انتقال ( آوند آبکش) در حال چرخش است. از بررسی دقیق مقادیر پتانسیل آب در این گونه بر می آید که حرکت رو به بالای آب در آوند آبکش تحت اثر شیب پتانسیل آب از منبع به مخزن صورت می گیرد این حرکات آب، قوانین ترمودینامیک را لغو نمی کند، چرا که حرکت آب بیشتر به کمک جریان جرمی و نه پتانسیل اسمزی، صورت می گیرد. ضمن انتقال مواد از یک لوله غربالی به لوله دیگر، هیچ غشایی قطع نمی شود و مواد نیز با سرعتی معادل سرعت آب حرکت می کنند. تحت چنین شرایطی، فشار اسمزی، گرچه پتانسیل آب را تحت تاثیر قرار می دهد، اما هیچ نقشی در ایجاد نیروی محرکه لازم برای حرکت آب ندارد. بنابراین حرکت آب در مسیر انتقال بیشتر براساس شیب فشار و نه شیب پتانسیل آب صورت می گیرد. البته بدون هیچ تردیدی انتقال فشاری غیرفعال در مسیرهای طولانی در لوله های غربالی تماماً به مکانیزمهای انتقال فعال دخیل در بارگیری و تخلیه آوند آبکش وابسته است. این مکانیزمهای فعال، مسئول برقراری شیب فشار در اولین نقطه هستند. پیشگوییهای از مدل جریان فشاری از آنجا که فرضیه جریان فشاری مورد قبول عموم است، باید در ارتباط با پیشگوییهای مبتنی بر این مدل، به برخی نکات توجه کرد اول آنکه منافذ عناصر غربالی نباید بسته باشند چرا که اگر پروتئین P یا هر ماده دیگر این منافذ را مسدود کند، مقاومت در برابر جریان عناصر غربالی به سرعت افزایش می یابد. دوم آنکه در یک عناصر غربالی واحد انتقال دو جهته واقعی ( یعنی انتقال در هر دو جهت) رخ نمی دهد. جریان توده ای محلول، مانع از انتقال دو جهته می شود، چرا که در یک لوله، محلول تنها می تواند در یک جهت حرکت کند! در آوند آبکش، حرکت دو جهته مواد محلول تنها در دستجات آوندی مختلف یا عناصر غربالی متفاوت میسر است سوم اینکه با توجه به فرضیه جریان فشاری که یکی از نظریات غیرفعال است صرف انرژی زیاد برای انتقال مواد در بافتهای مسیر لازم نیست. در این مسیر، انرژی تنها برای نگهداری ساختمان عناصر غربالی و سلامت غشای سلول لازم است بنابراین چنانچه در بافتهای مسیر، تیمارهایی نظیر درجه حرارت کم، اکسیژن کم و بازدارنده های متابولیکی که فراهمی ATP را کاهش می دهند اعمال شود. نباید انتقال متوقف شود. توجه داشته باشید که در ارتباط با وابستگی مکانیزم انتقال انرژی، دلیلی در دست نیست که مواد بازدارنده، انتقال را از طریق اضمحلال ساختمانی غشا و سلول و نه تنها از طریق کاهش فراهمی ATP متوقف می سازند. سلامت و کارایی غشای سلول برای حفظ مواد محلول در جریان انتقال و باز بودن منافذ صفحه غربالی برای جرکت رو به جلو این جریان لازم است. چهارم آنکه فرضیه جریان فشاری به وجود یک شیب فشار مثبت نیاز دارد بدین منظور باید فشار تورژسانس عناصر غربالی منبع، بیشتر از عنار غربالی مخزن باشد و اختلاف فشار نیز به اندازه باشد که بر مقاومت مسیر غلبه کرده و مواد را با سرعتی معادل آنچه اندازه گیری شده، حرکت دهد.
ممکن است مکانیزم انتقال در آوند آبکش بازدانگان با نهاندانگان متفاوت باشد سلولهای غربالی بازدانگان، از بسیاری جهات مشابه اجزای لوله های غربالی بازدانگان می باشند البته سطوح غربالی سلولهای غربالی نسبتاً غیرتخصصی هستند و به نظر نمی رسد که دارای منافذ باز باشند این منافذ به وسیله غشاهای متعددی که به شبکه آندوپلاسمی صاف مجاور سطوح غربالی متصلند، پر شده اند. قطعاً چنین منافذ با نیازهای فرضیه جریان فشاری تضاد دارد. به هر حال یا این تصاویر الکترونی با مشکلی در فرایند تثبیت همراه هستند و یا مکانیزم دیگری درانتقال بازدانگان ایفای نقش کرده است که شاید شبکه آندوپلاسمی در آن نقش مهمی دارد.
موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ چهار شنبه 4 مرداد 1391برچسب: مکانیزم انتقال در آوند آبکش, ] [ 11:48 ] [ حمیدرضا قاسمی ]
سلولهاي زنده آبكش، مواد غذايي مثل قندها را از طريق پلاسمادسماتا (Plasmodesmata) به ساير سلولها و اندامهاي گياه منتقل مي نمايند. بافت آبكش از عناصر آوند آبكش و سلولهاي همراه تشكيل شده است. آبكش از سلولهاي مادر لايه كامبيوم كه سلولهاي چوبي را نيز توليد مي كنند بوجود مي آيد. آبكش مانند چوب داراي الياف و سلولهاي پارانشيمي و سلولهاي شعاعي مي باشد. عناصر آوند آبكش شبيه عناصر چوبي از هر دو انتها بر هم منطبق بوده و لوله هاي آبكش را بوجود مي آورند. ولي بر خلاف عناصر چوبي ديواره انتهايي كاملا باز نبوده ولي در عوض پر از منفذهاي كوچكي هستندكه از درون آنها سيتوپلاسم از سلولي به سلول ديگر منتقل مي شود. اين مناطق منفذدار آوند آبكش، صفحات آبكش ناميده مي شوند. عناصر آبكش هنگام بلوغ بدون هسته مي باشند، ولي علارغم اين واقعيت سيتوپلاسم آنها براي انتقل مواد غذايي به صورت محلول در سراسر گياه بسيار فعال است. ظاهرا سلولهاي همراه ارتباط خيلي نزديكي با لوله هاي آبكش دارند و باعث انتقال غذا مي شوند. انتقال شیره پرورده براساس تئوری فشار اسمزی صورت می گیرد. ابتدا در منبع قندها به صورت فعال وارد آوندهای آبکش منبع شده سپس پتانسیل اسمزی در این آوندها افزایش یافته و آب مورد نیاز خود را از آوندهای چوبی گرفته و فشار افزایش یافته و مواد قندی و دیگر مواد موجود در آوند آبکش براساس شیب فشار به سمت مخزن حرکت کرده و در آنجا پس از تخلیه پتانسیل اسمزی مثبت تر شده و آب اضافی مجددا وارد آوند چوبی می شود. موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 30 تير 1391برچسب:مشخصات بافت آبکشي را به طور مختصر نوشته و مکانيسم حرکت مواد در آن را توضيح دهيد, , ] [ 18:6 ] [ حمیدرضا قاسمی ]
فضای آپوپلاستي به دو بخش فضای آزاد پخش (آب) و فضای آزاد دونان تقسیم می شود که در بخش اول مکانیسم حرکت یونهای معدنی صرفا شیب غلظت بوده و به تعادل می رسد و اگر ریشه در آب مقطر قرار گیرد تمام یونهای جذب شده به آب مقطر برمی گردد. اما در بخش دوم، یونه به ویژه کاتیونها جذب بارهای منفی ماکرومولکولهای تشکیل دهنده دیواره شده و تراکم آنها افزایش یافته و میزان جذب آنها افزایش می یابد. موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 30 تير 1391برچسب:انتقالات غيرفعال و قلمروهاي آنها را بنويسيد,, ] [ 18:5 ] [ حمیدرضا قاسمی ]
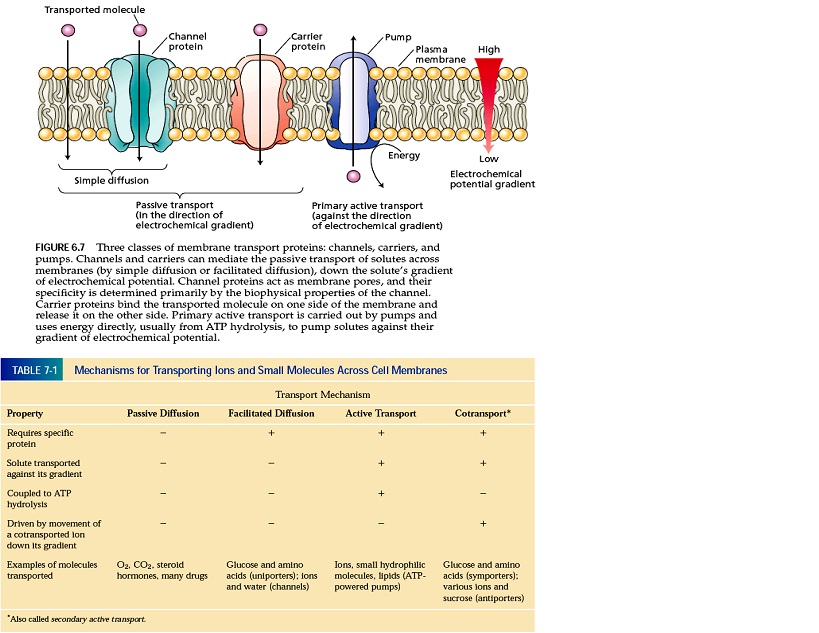
انتشار از خلال غشاء به صورت غیرفعال، در جهت شیب الکتروشیمیایی صورت گرفته و به دو بخش ساده و تسهیل شده تقسیم می شود. در انتشار تسهیل شده پروتئین های غشایی اعم از کانال ها (دایمرهای پروتئینی انتگرال غشایی با سرعت بالا 106 الی 108) و ناقل ها که در اثر تغییر شکل خود باعث انتقال مواد از خلال غشاء می شوند، نقش دارد. ناقل ها به عنوان یونی پورترها شناخته می شوند و دارای سرعتی برابر 102 الی 104 مولکول در ثانیه است. از کانال ها می توان به آکواپورین ها که حرکت آب را از غشاء سلول تسهیل می کند و این مسیر اول انتقال آب است و مسیر دوم، انتشار ساده از لابه لای غشای لیپیدی صورت می گیرد. از ناقل ها می توان به انتشار گلوکز و اسیدآمینه از عرض غشاء پلاسمایی سلول های پستانداران، اشاره کرد. موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 30 تير 1391برچسب:انتشار تسهيل شده چيست؟ و در چه سطوحي مي تواند عمل کند, با ذکر مثال آن را توضيح دهيد,, ] [ 18:2 ] [ حمیدرضا قاسمی ]
فضای آپوپلاستي به دو بخش فضای آزاد پخش (آب) و فضای آزاد دونان تقسیم می شود که در بخش اول مکانیسم حرکت یونهای معدنی صرفا شیب غلظت بوده و به تعادل می رسد و اگر ریشه در آب مقطر قرار گیرد تمام یونهای جذب شده به آب مقطر برمی گردد. اما در بخش دوم، یونه به ویژه کاتیونها جذب بارهای منفی ماکرومولکولهای تشکیل دهنده دیواره شده و تراکم آنها افزایش یافته و میزان جذب آنها افزایش می یابد. موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 30 تير 1391برچسب:انتقال آپوپلاستي يون هاي معدني وآب را در سطح ريشه توضيح داده و موانع اين انتقال را بنويسيد,, ] [ 18:0 ] [ حمیدرضا قاسمی ]
انتقال از عرض غشاء به دو صورت فعال (اوليه که به طور مستقيم وابسته به ATP، انتقال فعال ثانويه به طور غير مستقيم وابسته به ATP) و غير فعال (انتشار ساده وتسهيل شده به کمک پروتئين هاي غشائي اعم از کانال ها يا ناقل ها) صورت مي گيرد که در نوع دوم نيازي به مصرف انرژي نيازي نيست زيرا انتقال از غلظت بالاتر به غلظت پايين تر صورت مي گيرد. اما در نوع اول انتقال مواد، به مصرف انرژي نياز است زيرا مواد برخلاف شيب الکتروشيميايي (شيب غلظت و شيب الکتريکي) منتقل مي شود. منابع انرژي مورد نياز نيز به سه دسته تقسيم مي شود: الف- هيدروليز ATP ب- انرژي حاصل از واکنش هاي اکسيد و احياء نظير زنجيره انتقال الکترون در ميتوکندري و کلروپلاست. ج- گرفتن انرژي نور توسط مولکول ناقل مثل باکتريوردوپسين موجود در غشاء سلول هاي باکتريايي با جذب نور پروتون ها را از سيتوپلاسم خارج کرده و هنگام بازگشت اين پروتون ها يک نيروي مفيد (نيروي محرکه پروتوني يا PMF) براي ورود مواد موردنياز برخلاف شيب الکتروشيميايي (انتقال فعال ثانويه) مي تواند مورد استفاده قرارگيرد.
موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 30 تير 1391برچسب:منابع انرژي لازم براي انتقال غشايي چگونه تامين مي شود؟ , ] [ 17:40 ] [ حمیدرضا قاسمی ]
موضوعات مرتبط: کنکور، دانشگاه، اکولوژی، تکوین گیاهی، فیزیولوژِی گیاهی، سیستماتیک گیاهی، ، برچسبها: [ سه شنبه 2 خرداد 1391برچسب:دانلود رایگان سوالات دکتری نیمه متمرکز 91 , زیست شناسی, انگلیسی, استعداد تحصیلی, ] [ 12:19 ] [ حمیدرضا قاسمی ]
به معنای وسیع آن به هر عامل یا ترکیبی از عوامل محیطی اطلاق میگردد که باعث شود گیاه نتواند به اندازه توان بالقوه ژنتیکی خود رشد کند. به عبارت دیگر تنش عبارت است از هر گونه تغییر در عوامل طبیعی نسبت به شرایط بهینه رشد گیاه که رشد و نمو را کاهش و یا به طور نامطلوب تغییر داده و موجب کاهش رشد و عملکرد گیاه گردد. در بیشتر موارد تنش به عنوان دور شدن از شرایط معمول زندگی و ایجاد تغییرات و واکنشهایی در کلیه سطوح عملکرد در نظر گرفته شده است، این تغییرات قابل برگشت بوده و یا مکن است پایدار باقی بماند.
برای ادامه مطلب روی لینک زیر کلیک کنید: http://ecology.persianblog.ir/post/165/
موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: [ دو شنبه 28 فروردين 1391برچسب: تنش های محیطی, ] [ 11:25 ] [ حمیدرضا قاسمی ]
این سوال در چندین دوره ی آزمون دکتری مطرح شده و پاسخ آن فقط برای اعضاء قابل روئیت خواهد بود. لطفا به صورت رایگان عضو شوید.
موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ دو شنبه 28 فروردين 1391برچسب:چگونه پتاسیم (در سطح مولکولی) توسط ریشه گیاه جذب می شود؟, ] [ 11:19 ] [ حمیدرضا قاسمی ]
گونه هاي فعال اکسيژن و تنفس گياهي (Reactive Oxygen Species (ROS) and Plant Respiration) گونه هاي فعال اکسيژن نتيجه اجتناب ناپذير تنفس هوازي است. هنگامي که اکسيدازهاي انتهايي (سيتوکروم c اکسيداز و اکسيداز جايگزين) با اکسيژن واکنش مي کنند، چهار الکترون منتقل شده و آب توليد مي شود. در هرحال، گهگاه اکسيژن مي تواند با ديگر اجزاء انتقال الکترون واکنش کند. اما در اين حالت، فقط يک الکترون منتقل مي شود و به توليد آنيون سوپراکسيد (O2-) منجر مي شود. تخمين زده مي شود که در بافت هاي گياهي يک تا دو درصد اکسيژن مصرف شده به تشکيل آنيون سوپراکسيد منجر مي شود (1998Puntarulo et al,). در بين واکنش هاي متنوع ممکن، سوپراکسيد به پراکسيد هيدروژن، راديکال هاي هيدروکسيل و ديگر شکلهاي فعال اکسيژن که همه آنها مي توانند از راه هاي گوناگون باعث آسيب ديدگي و خسارت شوند، تبديل مي شود. زنجيره هاي جانبي پروتئين تغيير يافته و حتي محور اصلي ممکن است شکسته شود. به عنوان مثال، اسيدهاي چرب سيرنشده موجود در ساختمان ليپيدهاي غشاء مورد هجوم واقع شده، مشتقات پراکسيد تشکيل شده و سرانجام به شکسته شدن محور اصلي اسيد چرب ختم مي شود. در نهايت، شکلهاي فعال اکسيژن با مولکول DNA واکنش کرده و باعث جهش مي شوند. موضوعات مرتبط: تکوین گیاهی، فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ جمعه 18 فروردين 1391برچسب: گونه هاي فعال اکسيژن و تنفس گياهي (Reactive Oxygen Species (ROS) and Plant Respiration), ] [ 17:44 ] [ حمیدرضا قاسمی ]
رنگدانه ها و مواد آروماتيك در گلها و ميوه ها ،حشرات ، پرندگان و ديگر حيوانات را جذب ميكند كه گلها را تلقيح كرده يا ميوه ها را پراكنده كند ((zoochory. از آنجاببكه دو طرف شراكت (گياهان و جانوران) از اين رابطه سود مي برند (عمل جذب كنندگان بعنوان همزيست). رنگ گلها (گلبرگ، بساك،دانه گرده) ميوه ها از رسوب تركيبات رنگي در غشاهاي زيستي (كارتنوييد ها) و ديواره هاي سلولي (فلوبافنهاي قهوه اي ،ملانين سياه) و از رنگدانه هاي قند دار محلولدر خود رنگ به چندين عامل بستگي دارد: 1-نوع ساختار كروموفور(شكل 15-1)ساختار فلاون در فلاونوييدها ، ساختار بتايين دربتاسيانين و بتازانتين ها 2 - نوع جانشيني در ساختار پايه 3- كمپلكس ممكن باند شده به Fe+3 و AL+3 4 ـ pH شيره گياهي
به اين ترتيب درجات متفاوت رنگ بين دامنه اي از كرم،زرد ، زرد كمرنگ ، قرمز، آبي و ارغواني غليظ تغيير ميكند (جدول 5ـ1).نشانهاي شهد ماوراء بنفش بر روي گلها به وسيله همنوپتران و برخي از ديپترا (دوبالان) قابل تشخيص است. پرندگان با رنگهاي روشن و ماوراء بنفش منعكس شده توسط لايه هاي مومي سطح ميوه هاي خاصي جذب مي شوند .مواد آروماتيك حشرات ، پرندگان و پستانداران را به سوي گلها و ميوه ها دقيقاََ وقتي كه گلها باز ميشوند و ميوه ها مي رسند، جذب مي كند. انتشار عطر معمولاََ با فعاليتهاي روزانهَ حيوانات گرده افشان (پروانه ، زنبور) يا ميوه خواران هم زمان است (شكل 16-1).انتشار عطر يا بو به وسيله گياهان پيامي براي آمادگي جهت گرده افشاني است. بوهاي منتشر شده تقريباََ هميشه مخلوطي از متابوليت هاي ثانويه گياهي است به ويژه منوترپنها (روغنهاي اترال) الكلهاي زنجيري ، كتونها ، اترها ، اسيدهاي چرب، آروماتيكها (وانيلين،يوگنول) ،آمين ها و مشتقات ايندولها . يك مورد غير معمول گرده افشاني با جذب فوق العاده ويژه گرده افشانها توسط گونه هاي Ophrys وجود دارد. ترپنوييد هاي حلقوي ساطع شده از گلهاي اركيد فرومونهاي جنسي زنبورهاي وحشي را تقليد مي كنند.حركات جماع نرهاي گول خورده انتقالات پيچيدهَ دانه گرده به كلاله را با موفقيت فراهم ميكند. موضوعات مرتبط: اکولوژی، فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 18 فروردين 1391برچسب: جذب گرده افشانها و پراكند گي ميوه ها توسط حيوانات, ] [ 17:33 ] [ حمیدرضا قاسمی ]
گياهان مواد وترشحات متنوعي توليد مي كنند كه به طور منظم اثراتي را وقتيكه توسط ديگرگياهان،ميكرواورگانيسمها يا حيوانات جذب مي شوند،اعمال مي كنند. به طور طبيعي مواد بيو اكتيوي وجود دارند كه به عنوان ecomones يا Infochemicales شناخته مي شوند ونقشهاي مهمي در ايجاد انتقال پيامها،شناسايي،دفاع،ممانعت وتوليد سم بازي مي كند. برخي از اين عوامل فقط به صورت برون گونه اي به عنوان آتوتوكسين،ممانعت كننده در جوانه زني وبه عنوان فرومونها(جذب كننده وعلايم جاده اي)عمل مي كنند. درصورتيكه گروه ديگركه Allelochemicales ناميده مي شوند، برديگر گونه هاي گياهي،ميكرواورگانيسمها وحيوانات اثر مي كند. گروهي از اينفوكمكالها كه Allelomones ناميده مي شوند، فقط براي اورگانيسمهاي توليدكننده مفيد هستند،Kairomones گروه ديگري هستند كه فقط براي مصرف كنندهها مفيد هستند و Synomones براي دو طرف (شركا) مفيد هستند. طبيعت اغلب واكنشها نا معلوم ونسبتا پيچيده است،بنابراين مشكل است كه اثرات مواد بيواكتيورا بدون ابهام وصريح طبقه بندي كنيم. درك صحيح ما از واكنشهاي شيميواكولوژيكي براي فهم مااز فرايندهاي اكوسيستم وبراي پيشرفت اكولوژيكي دراندازه گيري درست بقا گياه لازم است.مواد بيواكتيو گياهي اغلب مواد واسطه يا محصولات نهايي متابوليسم ثانويه است. بنابراين آنها به عنوان مواد ثانويه گياهي معرفي مي شوند.آنها به صورت زيستي از پيشروهاي حاصله از متابوليسم اوليه سنتز شده اند. (شكل1-12)مهمترين راههاي سنتزي آنهايي هستند كهاز متابوليسم قند وچربي از طريق استيل كوانزيم آ،موالونات،ايزوپنتنيل پيروفسفات بهترپنوييد و استروييدها،از متابوليسم آمينواسيد و قند از طريق اسيد شيكميك وراه استات پلي كيتيد به تركيبات فنولي ومشتقاتشان(مثلافنيل پروپانها،فلاونوييدها،تاننها،موادگوناگون گلسنگ)واز طريق آمينو اسيدها به الكالوييدهامنتج مي شود. تخمين زده مي شود كه صد هزار مواد طبيعي اكوبيوشيميايي فعال وجود دارند. جدول(3-1) خلاصه اي از مهمترين گروههاي موادوانتشار(پراكندگي)تاكسونوميك آنهافراهم مي كند. تنوع موادثانويه گياهي درطول فيلوژني گياهي افزايش يافت ودر اثر فعل وانفعالات با پارازيتهاي ميكروبي وحيوانات مصرف كننده كامل شد. از آنجاييكه متخصصين مكررا درموردتمام انواع مواد دفاعي و توكسينهاي به توافق رسيده اند،تغييرات يا سنتز مواد جديد بايستي به طور مداوم پديدار شود. اين سازگاريهاي طبيعي در مدت نسبتا كوتاهي رخ مي دهد.اينها بديهي است كه پراكندگي شيميواكوتيپهاي متفاوت و اكوتيپهاي قابل تشخيص شيميوتاكسنومي(مثل تفاوتهاي بين عطر گونه هاي متفاوت لامياسه)قابل ملاحظه است. تركيبات عمومي فنيل پروپان (رزينها، ليگنين شكل 14-1)مشتقات فنولها (تاننهاي غليظ شده) و فلاونوييدهاي اولين مواد دفاعي شيميايي بودند كه استخراج شدهاند.(شكل13-1)طيف مواد طبيعي به ميزان زيادي با محصولات رده استات-موالونات افزايش يافت وقتي كه آنژيواسپرمها (نهاندانگان) تكامل يافتند و فرمهاي علفي پديدار شدند. كميت (اندازه)اينفوكمكالها در گياهان بين بافتها ، اندامها و حتي سلولهاي افراد متفاوت است. (عناصر ترشحي و تانن ايديوبلاستها) آنها در طول پيشرفت گياه و فصول متفاوت تغيير مي كنند. غلظت مواد بيوژنتيك گياهي ممكن است به ميزان قابل ملاحظه اي در طول روز يا حتي ساعت تغيير كند (نوسان كند). حالات غذايي گياه و اثرات ايجاد كنندهاي استرس متفاوت باعث تغيير قابل ملاحظه اي در كميت و تركيب (جدول 4-1) infochemical ها شده كه به نوبه خودفرايند هاي اكوسيستمها را تحت تاثير قرار مي دهد. حشرات بيشتر به گياهان استرس ديده و گياهاني كه به قدر كافي ذخيره نيتروژن دارند حمله مي كنند ، گياهان اساساًَ با افزايش توليد مواد دفاعي پاسخ مي دهند. روابط بيشمار از طريق انتقالات شيميايي بين ارگانيسم ها عمل مي كننذ. بنا بر اين تعدادي از وسيعترين و مهمترين پديده ها در زير توصيف ميشوند: موضوعات مرتبط: اکولوژی، فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 18 فروردين 1391برچسب: واكنشهاي شيميايي توسط مواد بيواكتيوگياه تغيير مي كند, ] [ 17:31 ] [ حمیدرضا قاسمی ]
ديواره گياهي به رسوباتي عمدتا پلي ساكاريدي اطلاق مي شود كه بر روي سطح بيروني غشاي سلولي قرارگرفته است. طبيعتا اولين سلول هايي كه ابتدا به وجود آمده اند، فاقد ديواره بوده اند و كسب ديواره در دوران تكامل اتفاق افتاده است. در دوران تكامل براي اولين بار، ديواره در باكتر ها شكل گرفته است. اين ها داراي ديواره هاي پپتيدوگليكاني بوده يعني از پروتئين و پلي ساكاريد ساخته شده و نحوه تشكيل ديواره در باكتري ها به صورت درون رو يا سانتري پت است. مواد وراثتي ابتدا دوبرابر شده، سپس بين دو سلول حاصله تقسيم مي شود. در باكتري ها تقسيم به صورت مستقيم (آميتوز) است. يعني ابتدا مواد وراثتي به نقطه معيني از ديواره متصل شده و پس از همانندسازي ديواره بين دو مولول وراثتي حاصله توسعه يافته و ديواره جديد دو مولكول وراثتي را از هم جدا مي كند. در خلال تكامل، هنگامي كه پروكاريوت ها به يوكاريوت ها تبديل مي شوند، موجوداتي نظير قارچ ها داريم كه يوكاريوت بوده ولي در آن ها ديواره به صورت سانتري پت شكل مي گيرد. قارچ هاي سيفومايست فاقد ديواره عرضي بوده و سيفوني هستند. اگر ديواره عرضي هم داشته باشند، سوراخ دار است. در قارچ هاي پيشرفته تر مثل آسكوميست ها كه هيف دارند، ديواره عرضي وجود داشته و سوراخ دار بوده كه اين سوراخ ها توسط اجسام پروتئيني (Plug) باز يا بسته مي شوند. ديواره سلولي در گياهان به صورت مركز گريز يا سانتريفيو ساخته شده و با توجه به گونه گياهي و رتبه تاكونوميكي آن متفاوت است. به طور كلي از سه لايه به نام هاي تيغه مياني، ديواره اوليه (كه در تمام سلوهاي گياهي وجود دارد) و ديواره ثانويه كه خاص سلول هاي تمايزيافته مثل آوندهاي چوبي است. البته در برخي سلول ها ديواره ديگري بر روي ديواره ثانويه به نام ديواره ثالثيه تشكيل مي شود. تيغه مياني بيشتر از پكتات كلسيم ساخته شده ولي ديواره اوليه از سلولز، همي سلولز و پروتئين هاي آنزيم و ساختماني مثل اكستنسين و اكسپنسين ساخته شده است و تعداد لايه هاي آن حداكثر چهار عدد مي باشد. تعداد لايه هاي ديواره ثانويه ممكن است به چهل عدد هم برسد. در هر لايه سه تركيب ديواره اوليه وجود داشته ولي مقدار سلولز ان بيشتر بوده و رشته هاي سلولزي از نظم بيشتري برخوردارند. همچنين اين رشته در هر لايه موازي همديگر بوده ولي با رشته هاي لايه مجاور خود زاويه 120 درجه مي سازند. موضوعات مرتبط: تکوین گیاهی، فیزیولوژِی گیاهی، ، برچسبها: [ جمعه 18 فروردين 1391برچسب: تکامل ديواره سلولي را در بين موجودات زنده توضيح دهيد,, ] [ 17:24 ] [ حمیدرضا قاسمی ]
مترجم : حمیدرضا قاسمی موضوعات مرتبط: تکوین گیاهی، فیزیولوژِی گیاهی، ، برچسبها: [ دو شنبه 14 فروردين 1391برچسب: آنزیم ها و عواملی دیگر که انعطاف پذیری دیواره سلولی را افزایش می دهد , ] [ 13:20 ] [ حمیدرضا قاسمی ]
خلاصه واکنش های نوری کتاب تایز: - انرژی نور جهت حرکت الکترونها از طریق گیرنده ها و دهنده ها و همچنین انتقال H+ در عرض تیلاکوئید و تولید نیروی محرکه پروتونی ((PMF استفاده می شود. - طیف جذبی: میزان انرژی جذب شده توسط یک مولکول یا ماده بعنوان تابعی از طول موج نور را نشان می دهد. - مولکول کلروفیل پس از دریافت نور قرمز و آبی از حالت بنیادی خارج و به حالت بر انگیخته منتقل می شود هر چه انرژی فتون دریافتی بیشتر باشد، بر انگیختگی شدیدتر و مدّت زمان آن کمتر خواهد بود و پایدار ترین حالت برانگیختۀ خود می رسد. از این حالت تا رسیدن به وضعیت پایه به صورت های متفاوتی انرژی تلف می شود:1- فلولؤرسانس 2- تبدیل مستقیم به گرما 3- ترافرستی یا انتقال انرژی به مولکولدیگر 4- واکنش فتوشیمیایی موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ دو شنبه 14 فروردين 1391برچسب: خلاصه واکنش های نوری کتاب تایز, ] [ 13:15 ] [ حمیدرضا قاسمی ]
جذب در گیاهان مقدمه: مکانیسم جذب یون ها توسط سلول های گیاهی طی سال های اخیر مورد مطالعات دقیق قرار گرفته است. علی رغم آنکه ارتباط یونی بین سلول های گیاهی با محیط اطراف از چندین نظر با سلول های جانوری اختلاف دارد، اما به طور کلی می توان از مطالب مربوط به الکتروفیزیولوژی سلول های جانوری و ویژگی های سلول های عصبی، برای سلول های گیاهی نیز استفاده نمود. از طرف دیگر کوچک بودن اندازه سلول های گیاهی، کار مطالعه آن ها را مشکل می سازد. بنابراین دانشمندان اغلب در مطالعات خود از سلول های بزرگ جلبک های مختلف که دارای هستۀ مرکزی می باشند، استفاده می نمایند و شاید به همین دلیل اطلاعات در مورد جلبک ها از گیاهان بیشتر است. منبع: http://microbiology-sh.blogfa.com/post-140.aspx موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ شنبه 12 فروردين 1391برچسب:جذب یون ها در گیاهان, ] [ 16:38 ] [ حمیدرضا قاسمی ]
این سوالات مربوط به 5 سال گذشته دانشگاه آزاد است موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب
تهيه شده توسط خانم مهندس ندا زرگاني موضوعات مرتبط: فیزیولوژِی گیاهی، ، برچسبها: ادامه مطلب [ شنبه 12 فروردين 1391برچسب:اتيلن, ] [ 16:13 ] [ حمیدرضا قاسمی ]
|
درباره وبلاگ  کارشناس علوم گیاهی، کارشناس ارشد بوم شناسی گیاهی و دانشجوی دکتری فیزیولوژی گیاهی دانشگاه اصفهان
|
| [ طراحی : ایران اسکین ] [ Weblog Themes By : iran skin ] | ||
 آمار
وب سایت:
آمار
وب سایت: